



Developing Understanding Through Model-based Inquiry
James Stewart
Jennifer L. Cartier
Cynthia M. Passmore
Department of Curriculum & Instruction,
University of Wisconsin-Madison
National Center for Improving Student Learning & Achievement in Mathematics
and Science
Developing Understanding Through Model-based Inquiry
| Understanding in science includes being familiar with a discipline's concepts, theories, and models, an appreciation for how knowledge is generated and justified, and the ability to use such knowledge to engage in inquiry. |
Teaching for, and learning with understanding are desirable educational goals and have been the centerpiece of two significant national reform efforts in science education (NRC, 1996; AAAS, 1993). The positions in the National Research Council and the American Association for the Advancement of Science documents reflect the view that understanding involves a set of interrelated components that form the basis for knowing about and interacting with the natural world. These components includes a familiarity with a discipline's concepts, theories, and models, an understanding of how knowledge is generated and justified in various disciplines, and the ability to use such understandings to engage in inquiry.
The view taken in this chapter is that scientific practice begins with the recognition (mediated by the conceptual lenses provided by appropriate explanatory models) of some phenomena warranting explanation. From data about natural phenomena, scientists then must establish patterns related to the data and then create, revise, or use explanatory models to account for these data patterns. Clearly, data collection, pattern recognition, and proposing, revising, or defending explanatory models in science is, in reality, both a highly discipline-specific process and a seamlessly integrated one; that is, data collection and analysis proceeds concurrently with the use and production of explanatory models. However, for education purposes, the generalized process describe above has proven to be quite powerful. While disciplinary constraints dictate just what constitutes a phenomenon and what types of patterns are sought we have found that this framework can be useful for identifying important phenomena, patterns and models in various disciplines and developing curricula around these central ideas. The specificity of disciplinary practice comes into play in such curricula when students begin to discuss and defend their models with one another. The structure of their explanatory models, as well as the body of scientific ideas in relation to which their models are judged, is discipline-specific.
| In this chapter we illustrate how high school students have come to understand sophisticated science concepts by participating in model-based inquiry. |
Given the importance of models and modeling in science classrooms (and in science), it is essential that students learn about scientific models (in addition to learning the concepts relevant to particular models) and how to communicate with one another regarding the use and evaluation of such models. In this chapter we will illustrate how high school students have been able learn about science as model-based inquiry generally while also coming to understand significant discipline-specific concepts and inquiry procedures through participation in inquiry in genetics and evolution. The introduction to the chapter begins with a classroom vignette. Interestingly the vignette-while it portrays a classroom in which students are generating data, seeking patterns, and proposing and revising explanatory models-does not showcase students engaged in learning new scientific concepts. Don't be alarmed! The vignette illustrates important aspects of a model-based inquiry classroom : such "science-free" modeling activities are used to introduce students to science as a modeling game, to develop classroom norms for interacting with other students, and to establish scientific norms of argumentation concerning data, claims, explanatory models, and their relationship. Following similar introductory activities students then engage in similar intellectual pursuits in the context of sophisticated disciplinary content.
An Initial Modeling Experience
| A class of high school students is examining a half-gallon milk-carton shaped container that pours a pre-measured amount of liquid laundry soap each time it is tipped. They are engaged in what they consider to be a central activity of scientists-making observations.
The students respond with a variety of observations.
After several minutes of listening to the students, their teacher stops them and invites them to take a closer look at the carton, prompting them to identify patterns associated with their observations. Their reflection on these patterns leads the students to propose manipulations of the container-these in turn produce more observations. The teacher now interrupts them to comment on the high quality of their work, saying,
She goes on to challenge them,
Over the next two class periods, they work in animated groups to develop models that can account for their observations. They describe, draw, and build three-dimensional representations of what they think is in the carton. They argue. They negotiate. They revise. Then they share drawings of their models with one another.
The students also propose tests of their models.
A visitor to the classroom would notice that Mrs. S. listens attentively to the descriptions that each group gives of it's model and the observations that the model is designed to explain. She pays special attention to their interactions with other groups, and is skillful in how she converses with them during their presentations. Through her comments she demonstrates how to question the models of others and how to present a scientific argument. To one group she says: "I think I follow your model, but I am not sure how it explains why you get 90 milliliters of liquid each time you tip the box." To another she comments, "You say that you have used something similar to a toilet bowl valve. But I don't understand how your valve allows soap to flow in both directions." And to a third group she asks, "Do you think that Ian's model explains the data? What question would you ask his group at this point?" By the end of the three-day activity the students are explicit about how their prior knowledge and experiences influence their observations and their models. They also ask others to explain how a proposed model is consistent with the data, and challenge them when a component of a model, designed to explain patterns in observations, doesn't seem to work as it has been described. |
Curriculum Design Principles
Introduction
| We consider a scientific model to be an idea or set of ideas that explains the causes of particular natural phenomena. Models are complex constructions that consist of conceptual objects (e.g., alleles, populations) and processes (e.g., selection independent assortment) in which the objects participate or interact. Hafner & Culp, 1991 |
Over the course of several days, the students in this classroom have experiences that, in important respects, are similar to those of scientists, including:
- using prior knowledge to pose problems and generate data.
- searching for patterns in data.
- developing scientific (or explanatory) models to account for patterns.
- using patterns in data and their models to make predictions.
- revising initial models in light of anomalous data and in response to critiques of others.
- making their ideas and their thinking public.
In short, they are participating in a classroom scientific community. Such participation is a hallmark of classrooms where model-based inquiry curricula are used. In this chapter we will describe three design principles that are central to developing such curricula and the ways in which these principles have been implemented in high school instruction in genetics and evolutionary biology. The core design principles, consistent with three major themes of the National Research Council Book How People Learn (NRC, 1999), are:
| We encourage readers to visit the web site (http://ncisla.wceruw.org/muse/) that serves as a companion to this chapter. For each of our multi-week curricula, the site includes—
|
Attend to Significant Disciplinary Knowledge-such knowledge includes the conceptual structures (concepts and explanatory models) of a discipline as well as how such structures are developed and the role they play in inquiry.
Attend to Student Knowledge-students may have significant prior knowledge, including misconceptions, related to scientific inquiry and science concepts. Their prior knowledge needs to be considered when developing curricula and related instructional materials.
Engage Students in Reflective Scientific Practice-classrooms should be places where all students are full participants in learning science by participating in realistic inquiry. Moreover, such participation leads to rich opportunities to reflect and learn about knowledge development in science.
This particular approach to curriculum development has emerged as a result of collaborative work with high school teachers and their students (our collaborative group is known as MUSE, or Modeling for Understanding in Science Education) As a part of that collaboration we have conducted research on student learning, problem solving, and reasoning. This research has led to refinements in the instruction, which in turn have led to improved student understanding. The remainder of this chapter is organized so that Section One provides an overview of the three design principles, while Sections Two and Three contain descriptions of these principles as they are instantiated in MUSE genetics and evolution curricula. Section Four describes some common elements in both of these classrooms that enable students to participate in reflective scientific practice.
Section One: An Overview of the Design Principles
Attending to Significant Disciplinary Knowledge
| Disciplinary knowledge and ideas about the nature of scientific inquiry are instantiated in classrooms where students learn by "doing science." |
For well over a decade we have developed science curricula where both disciplinary knowledge and knowledge about the nature of science comprise the student learning outcomes. Such learning outcomes are realized in classrooms where students learn by "doing science" in ways that are similar to the work that scientists do in their intellectual communities. We have created classrooms in which students are engaged in discipline-specific inquiry (empirical and conceptual investigations) as they learn and employ the explanatory models and the reasoning patterns of the discipline. In the descriptions of the genetics and evolution courses that follow it will become apparent that, while explanatory models are central in both disciplines, there are very different reasoning patterns involved in the use or construction of such models. This comes about largely because a primary activity in the practice of evolution, the reconstruction of past events, is not common in the practice of genetics.
Attending to Student Knowledge
| Understanding students' prior knowledge has influenced our work in the areas of—
|
Our work has benefited from the general research on student knowledge (outlined in Chapter Z). However, we have benefited even more from research about student learning in the fields of genetics and evolution and research about students' knowledge of aspects of the "nature of science" (especially that which speaks directly to models and modeling). An understanding of student prior knowledge has had a significant influence on our decision making in three areas:
- curriculum (selecting and sequencing student learning outcomes);
- instruction (designing and selecting tasks and activities for students); and
- assessment (determining the extent to which students have realized the intended learning outcomes).
Curriculum (Selecting and Sequencing Learning Outcomes): The primary basis for selecting learning outcomes is the practice of the appropriate discipline. That is, each discipline has at its core a set of key explanatory models, assumptions, and particular argument structures. These form the basis of MUSE curricula in various disciplines. Even so, the choice of learning outcomes is informed by the literature on student understanding of specific disciplinary concepts and from our knowledge of student performance in previous versions of the genetics and evolution courses. For example, the evolution education literature has made it clear to us that many students: (1) believe that evolution results from teleological processes; (2) do not consider evolution to be truly scientific (it lacks experimentation and directly observable events); and, (3) that science need not be limited to naturalistic, mechanistic explanations (see for example, Demastes, Trowbridge, & Cummins, 1992; Jensen, & Finley, 1995; Rudolph, & Stewart, 1998; Wirtz, 1993). With these understandings in mind we have focused upon the metaphysical, methodological, and epistemological perspectives of students and scientists in addition to the conceptual components of evolutionary biology as we developed the model-based inquiry evolution curricula.
| Students' Prior Knowledge
• Many high school students do not understand the interrelationship
of genetic, meiotic, and bio-molecular models, relationships that
are key to a deep understanding of inheritance phenomena. • Even after instruction, students may still believe that
mutations or "abnormal" genes are recessive, and that
dominant traits are more frequent in the population and more likely
to be inherited and/or selected for. • Students misunderstand the primacy of naturalistic explanations
in science. • Students have difficulty in understanding the concept of
variation that is a foundation of the natural selection model. • Students have misunderstanding about the origin, function,
and very nature of scientific models. |
While the selection of learning outcomes is informed by research on student prior knowledge we have found that it is equally important to take that research into consideration when sequencing those learning outcomes. For example, many high school students do not understand the interrelationship of genetic, meiotic, and bio-molecular models, relationships that are key to a deep understanding of inheritance phenomena (Cartier, 2000a; Kindfield, 1994; Wynne, Stewart, & Passmore, in press). To offset this problem we have identified learning outcomes that address the conceptual connections among families of models at the cytological, genetic, and bio-molecular levels and to introduce these models in a sequence that emphasizes their relatedness.
In the evolution course, knowing that students misunderstand the primacy of naturalistic explanations, we designed experiences that require students to identify the assumptions about mechanism (not all naturalistic) contained in the writings of William Paley, Jean Baptiste Lamarck, and Charles Darwin. Through these activities, which occur early in the instruction, students have opportunities to consider and debate the reasons why scientists feel it is important to limit themselves to naturalistic explanations. From this point on, we are able to focus only on Darwin's naturalistic idea of evolution by natural selection. In addition, this introductory comparison also allows us to address learning outcomes designed to eliminate teleological explanations (evident in Lamarckian-like explanations that students give for evolutionary phenomena) that have been widely reported in the literature (see for example, Clough & Wood-Robinson, 1985a, 1985b; Jensen & Finley, 1996). Details of how understanding student prior knowledge of the conceptual content of particular disciplines, about models and modeling, and about what it means to learn science will be returned to in Sections Two and Three.
Instruction (Development of Student Tasks and Classroom Structure): An awareness of typical misconceptions and difficulties that students may experience when learning particular content is important to the development of instructional tasks. Throughout our work we have carefully designed instructional activities that address students' prior knowledge and misconceptions related to understanding in genetics and evolutionary biology.
In addition to our concerns with student knowledge in particular content areas we have also found, as have others (Grosslight, Unger, Jay, & Smith, 1991), that students have misunderstandings about the origin, function, and very nature of scientific models. Because we expect students to develop understanding of the nature of models and of appropriate norms of argumentation in modeling classrooms we have developed introductory activities to address "nature of science" learning outcomes explicitly. For now it is enough to say that we begin instruction with a "science free" activity. For example, the genetics instruction begins with a black box activity (similar to the detergent-container described in the vignette that opened the chapter) that introduces students to the nature of explanatory models—they are ideas used to guide investigations. Similarly, the evolution course begins with a cartoon sequencing activity that provides students with a sense of what is involved in reconstructing the past (a central goal of inquiry in evolutionary biology). Both of these activities serve a common purpose-to introduce students to important ideas about the nature of knowledge generation and justification in particular disciplines.
| Because we expect students to develop understanding of the nature of models and of appropriate norms of argumentation in modeling classrooms, we have developed introductory activities to address "nature of science" learning outcomes explicitly. |
| By having tasks that require students to attend to knowledge across domains and by structuring classrooms so that students have to make their thinking about such integration public we have seen improvements in their understanding of genetics. |
Of course, there are many instances in our curriculum development work where an understanding of student conceptual knowledge related to a particular discipline is influential. For example, in early versions of the genetics instruction, it became apparent that students were solving problems, even sophisticated ones, without adequately drawing on an integrated understanding of meiotic and genetic models (Cartier, 2000a, Wynne, Stewart & Passmore, in press). In response, we designed a set of data analysis activities and related homework activities that required students to integrate across cytology, genetics, and molecular biology when conducting their genetic investigations and when presenting model-based explanations to account for patterns in data that they had generated. By having tasks that require students to attend to knowledge across domains and by structuring classrooms so that students have to make their thinking about such integration public we have seen improvements in their understanding of genetics (Cartier, 2000b).
| Knowing that students often perceive evolution to be an individual rather than a population-level process can provide teachers with an ear for listening to student discussions. We have seen, time and again, teachers who, when they become aware of the common struggles of students, begin to "hear" their own students differently. |
Assessment: Assessing the extent to which students realize intended learning outcomes needs to be undertaken with an eye to the various types of prior knowledge just described (misconceptions of science concepts, ideas about what science is, and the extent to which students' knowledge is integrated). It is fairly obvious how, at least in general terms, this might be accomplished. The identification of prevalent student knowledge, including misconceptions, can influence both formal and informal assessment activities. For example, knowing that students often perceive evolution to be an individual organism rather than a population-level process can guide the development of exam questions as well as provide teachers with an ear for listening to student discussions. We have seen, time and again, teachers who, when they become aware of common struggles of students, begin to "hear" their own students differently. Similarly, in genetics, test questions, or questions of students as they work in research groups solving problems can be structured to provide evidence of their understanding of the relationship of genetic and meiotic models. Thus, an important feature of instructional activities that provide students with opportunities to make their thinking and knowledge public, and therefore "visible" to teachers, is that they make assessment and instruction seamless. This becomes possible when students articulate the process of arriving at solutions and not simply the solution itself.
It is clear that knowledge of student understanding can be influential in the informal and formal assessment work of teachers. However, we believe that students also benefit from assessments that provide them with opportunities to see how their understanding may have changed over the duration of some unit of study. One method that we have found to have such potential is to require each student to critique her own early work based on what she knows at the conclusion of a course. Not only does this approach provide teachers with insights into student knowledge but it provides students with a glimpse at how much their knowledge, and their ability to critique arguments, has changed.
Promoting Reflective Scientific Practice
| Reflective practice involves an understanding of central explanatory models, the ability to use such models to conduct inquiry, and an understanding of and ability to engage in the assessment of scientific models. |
By considering authoritative accounts of scientific practice and research on students' knowledge it is possible to develop instruction that enable students to engage reflectively in scientific practice. Reflective practice involves an understanding of central explanatory models, the ability to use such models to conduct inquiry, and an understanding of and ability to engage in the assessment of scientific models. We have found that these outcomes can be realized in classrooms where students are full participants in a "scientific community" (Cartier, 2000b, Passmore & Stewart, 2000) Interestingly, one unexpected outcome of structuring classrooms so that students are expected to participate in the "intellectual work" of science has been an increased involvement and achievement by students who had not previously been identified as successful in science. Creating contexts for reflective practice will be illustrated in the descriptions of the genetics and evolution classrooms and will also be returned to in the conclusion to the chapter.
| For students to understand science content as well as what it means to make and justify claims in modeling classrooms, they need to be placed in situations where they are expected to think about their own thinking. |
To conclude Section One, we repeat that in order for students' understanding of science content and of how to make and justify claims in modeling classrooms to progress, they need to be placed in situations where they are expected to think about their own scientific thinking. Each of the MUSE classrooms has been carefully constructed to provide just such opportunities. Sections Two and Three include detailed illustrations of how this occurs in genetics and evolutionary biology.
Section Two: Developing Models of Inheritance in High School Genetics
| Student Inquiry in Genetics Nineteen students are sitting at desks and lab tables in a small and cluttered high school biology classroom. The demonstration desk at the front of the room is barely visible under the stacks of papers and replicas of mitotic cells. A human skeleton wearing a lab coat and a sign labeled "Mr. Stempe" stands in a corner at the front of the room and the countertops are stacked with books, dissecting trays, and cages holding snakes and gerbils. During the previous few days the students in this class studied the work of Gregor Mendel, the nineteenth century monk who has come to be known as the "Father of Genetics." Mendel grew generation after generation of pea plants in an attempt to understand how certain traits were passed from parent plants to their offspring. Years of Mendel's work resulted in the publication of Experiments on Plant Hybridization, (1865), a paper in which Mendel put forth his model explaining inheritance of discrete traits in plants1. The students read an edited version of this paper and refer to Mendel's idea as the "simple dominance model" because it explains the inheritance of traits derived from two alleles (or pieces of genetic information) when one of the alleles is completely dominant to the other. (see Figure 1A) During class on this day, the students' attention is drawn to the cabinet doors along the length of the room. These doors are covered with student drawings of family pedigrees labeled Summers: Marfan (See Figure 2), Healey: Blood Types, Jacques: Osteogenesis Imperfecta, and Cohen: Achondroplasia. The teacher is standing at the side of the room facilitating a discussion about these family pedigrees.
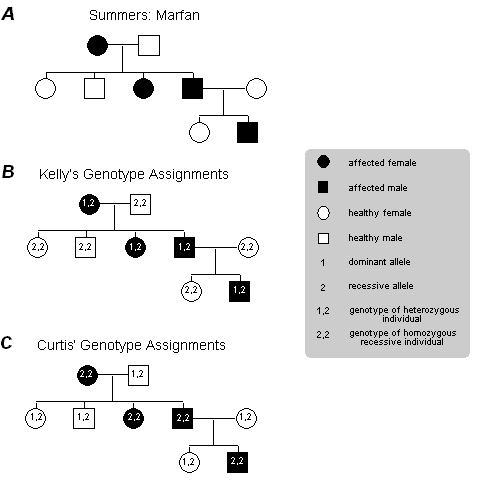
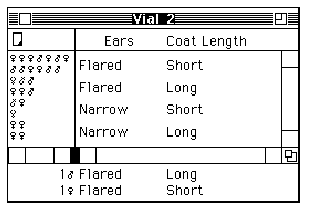
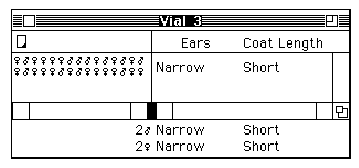
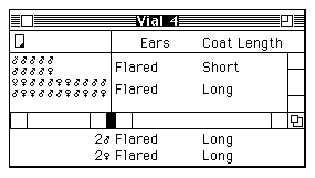
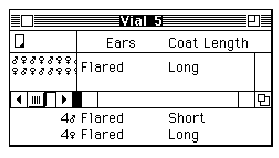
Kelly pulls a stool over to one of the cabinets at the side of the room, climbs up, and begins to label each of the circles and squares on the pedigree with two alleles: Some are assigned the genotype 1,2 (heterozygous, or possessing two different alleles) and others 2,2 (homozygous recessive, or possessing two recessive alleles) (see Figures 2A and 2B).
Curtis proceeds to label the same pedigree consistent with his idea that the Marfan allele is actually recessive (see Figure 2C).
|
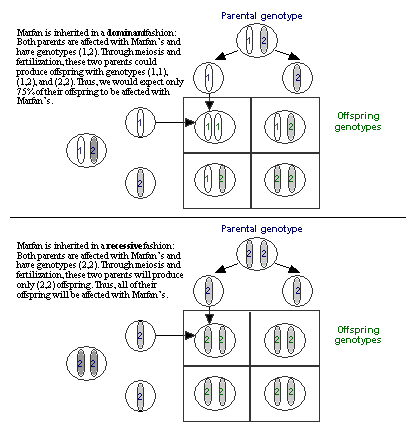
Figure 1 |
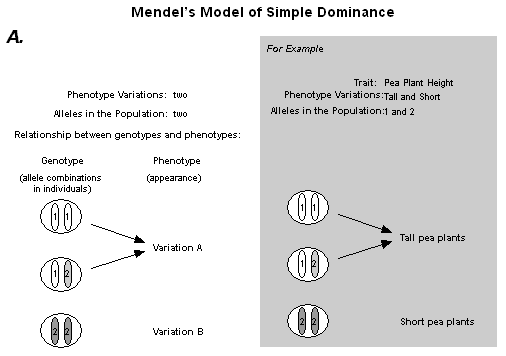
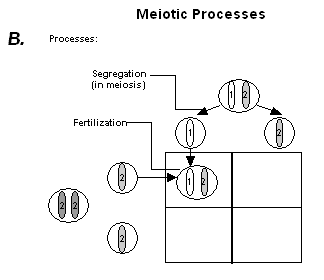
Figure 2
Figure 2: Pedigrees representing inheritance of Marfan Syndrome in the Summers family. (A) The original Pedigree represented in the inheritance pattern within the Summers family without specifying individual genotypes. (B) Kelly's genotype assignments, assuming that Marfan Syndrome is inherited as a dominant trait. (C) Curtis' genotype assignments, assuming that Marfan Syndrome is inherited as a recessive trait. |
The students in the high school biology class that we just 'peeked into' are engaged in genetic inquiry: They are examining data and identifying patterns of inheritance for various traits. They are also attempting to use a powerful explanatory model, Mendel's model of simple dominance, to account for the patterns that they see. And just as practicing scientists do, they recognize the limitations of their model when it simply cannot explain certain data patterns. These students are poised to continue their inquiry in genetics by revising Mendel's model such that the resulting 'new' models will be able to explain a variety of inheritance patterns, including the multiple allele/codominance pattern within the Healey pedigree.
Attending to Significant Disciplinary Knowledge
| The genetics unit in this biology class reflects important aspects of authentic genetic practice, particularly the use of essential explanatory models to account for and predict complex inheritance patterns in organisms. |
The genetics unit in this biology class reflects important aspects of authentic genetic practice, particularly the use of essential explanatory models to account for and predict complex inheritance patterns in organisms. We have already discussed the basis for emphasizing in our curricula particular ideas about the nature of science-these include how and why scientists use models to explain natural phenomena and the general criteria they apply when judging the utility of such models. In this section, we will focus on the genetics concepts that are foundational in our biology unit and how we engage students in model assessment and revision in this context.
| In choosing which genetics concepts to place at the core of our curriculum, we attempted to identify a set of important disciplinary concepts that would provide a rich context for illustrating how those concepts were generated and justified through inquiry. |
The choice of which genetics concepts to place at the core of our curriculum was governed by our belief that whatever specific subject matter we chose should not only be central to understanding in the discipline of genetics, but should also lend itself to be studied from the perspective of model-based inquiry; that is, we wanted a set of important disciplinary concepts that would provide a rich context for illustrating how those concepts were generated and justified. Thus, we selected the set of concepts associated with an understanding of classical Mendelian inheritance as a foundation to our genetics unit. In fact, the development of modern genetic theory from its classical Mendelian origins has been the subject of much historical and philosophical analysis. In Theory Change in Science (1991), philosopher Lindley Darden draws on historical evidence to identify a set of strategies scientists utilized to generate and test ideas while conducting early inquiries into the phenomenon of inheritance. She traces the development of a number of inheritance models that were at least originally seen to be at odds with those underlying a Mendelian (i.e. simple dominance) explanation of inheritance. Among these are the notions of linkage and multiple forms (alleles) of a single gene. In short, Darden provides a history of model-based inquiry into the phenomenon of inheritance-from a classical genetics perspective. Drawing on Darden's work and our own experiences as teachers and researchers in genetics, we were able to see how engaging students in building and revising Mendel's simple dominance model could provide rich opportunities to learn important genetics concepts as well as key ideas about the practice of genetics.
| Early instruction in the genetics class includes a few days during which students learn about the meiotic model and what phenomena this model can explain. |
Early instruction in the genetics class includes a few days during which students learn about the meiotic model and what phenomena this model can explain. This instruction will be discussed in more detail below in the section about Attending to Student Knowledge and is described only briefly here. However, we wish to point out that, although Mendel did not have the advantage of a basic meiotic model with which to begin his work, it has proven fruitful to start here with students. In an introductory activity, students look at sets of pictures and are asked to determine which are members of the same families. The bases for their decisions include physical similarities between parents and children and between siblings. Thus, instruction about meiosis focuses on how the meiotic model can account for these patterns: Children resemble their parents because they receive information from both of them; and siblings resemble each other but are not exactly alike because of the random assortment of parental information in meiosis.
| Meiosis is the process by which sperm and egg cells are formed. During meiosis, chromosomal replication is followed by two rounds of cell division. Thus, one cell undergoing meiosis produces four new cells each of which contain half the number of chromosomes of the original parent cell. See figure 1B for a representation of meiotic processes that underlie
Mendel's simple dominance model of inheritance. |
After students have developed some understanding of meiosis, they "create", with guidance from the teacher, a representation of Mendel's model of simple dominance (see Figure 1A) in an attempt to further explain why offspring look like parents. First, "Mendel" (a teacher dressed in a monk's robe and wearing antique spectacles) pays the class a visit and tells them that he'd like to share some phenomena and one important model from his own research with them. In character, "Mendel" passes out three packets of peas representing a parental generation and the F1 and F2 generations (the first and second filial generations, respectively). He asks students to characterize the peas according to color and shape. For example, the parental generation includes a round green pea and a wrinkled yellow pea. The F1 generation contains only round yellow peas. Finally, the F2 generation contains a mixture of round yellow, wrinkled yellow, round green, and wrinkled green peas in a ratio of approximately 9:3:3:1. Using what they already know about meiosis-particularly the fact that offspring receive information from both parents-they reconstruct Mendel's model of simple dominance to account for these patterns (see Figure 1A).
| After students have developed some understanding of meiosis, they "create", with guidance from the teacher, a representation of Mendel's model of simple dominance (see Figure 1A) |
While Darden's work aided in the identification of important inheritance models and strategies scientists used to judge those models, it was the work of philosopher Philip Kitcher (1984, 1993) that put the simple dominance model developed by students into context with comparable models of geneticists. According to Kitcher, genetic 'schemata'-which we take to be analogous to explanatory 'models'-provide the following information:
| The genetic models of both scientists and students provide information about different alleles for a given trait and the ways in which they combine to give rise to particular phenotypic variants. |
(a) Specification of the number of relevant loci and the number of alleles at each locus; (b) Specification of the relationships between genotypes and phenotypes; (c) Specification of the relations between genes and chromosomes, of facts about the transmission of chromosomes to gametes (for example, resolution of the question whether there is disruption of normal segregation) and about the details of zygote formation; (d) Assignment of genotypes to individuals in the pedigree. (1984, p.356)
Moreover, Kitcher describes how such schemata (models) might be used in inquiry:
[A]fter showing that the genetic hypothesis is consistent with the data and constraints of the problem, the principles of cytology and the laws of probability are used to compute expected distributions of phenotypes from crosses. The expected distributions are then compared with those assigned in part (d) of the genetic hypothesis. (p.356)
With their teacher's guidance, students represent the simple dominance model of Mendel in a way consistent with Kitcher's description of the models of geneticists. They pay particular attention to (b) and (d) above: Specifying the relationships between genotypes and phenotypes and identifying the genotypes of individuals in their experimental populations. Because our unit doesn't address multi-gene traits, one locus per trait is assumed (thus part of criterion (a) above is not applicable in this case) and students focus on determining the number of alleles at that locus. Finally, the students' prior understanding of meiosis-developed earlier in the unit-enables them to specify chromosomal transmission of genes for each particular case (letter (c) above). A representation of Mendel's model of simple dominance is shown in Figure 1A.
| The software program GENETICS CONSTRUCTION KIT or GCK (Calley & Jungck, 1997) enables students to study populations of "virtual fruit flies," noting patterns in the inheritance of various traits such as eye color, wing shape, etc. Students use their genetic models to explain data that they generate using GCK as well as other types of data pertaining to the inheritance of human diseases. |
Once the students have represented and used the simple dominance model to explain some phenomena, such as inheritance of characteristics in peas and disease traits in humans, the students use the model to explain data that they generate using the software program genetics construction kit or GCK (Calley & Jungck, 1997). This program enables students to manipulate populations of virtual organisms (usually fruit flies) by performing matings (or crosses) on any individuals selected. Each cross produces a new generation of organisms whose variations for particular traits are described. Thus, the students develop expertise using the simple dominance model to account for new data and they also design and perform crosses to test their initial genotype to phenotype mappings within these populations. Figure 3 is an excerpt from one student's work with GCK and the simple dominance model.
Figure 3
|

| The next step is to revise Mendel's simple dominance model to account for anomalous inheritance patterns such as—
|
Next, the teacher presents or revisits phenomena that the simple dominance model cannot explain. For example, students realize during the application of the simple dominance model to explain their human pedigrees that it is inadequate in some cases: It can't be used to account for inheritance of human blood types or achondroplasia. The next step for the class is to study these "anomalous" inheritance patterns using GCK. They begin with achondroplasia, a trait for which there are three variations rather than two. Students revise the simple dominance model to account for the codominant inheritance pattern observed for this trait. While solving GCK problems such as this one, students propose models that specify some or all of the information (a through d) listed above and then test their models for fit with existing data as well as for the ability to accurately predict the results of new experiments.
| Geneticists must assess a new inheritance model in part based upon how well it fits within an existing family of related models such as meiosis (including cytological data) and molecular biology (which specifies the relationships between DNA and proteins as well as protein actions in cells). Students must also judge their models based upon their consistency with previous models or ideas. |
Since most students ultimately explain the inheritance of achondroplasia using a codominant model (one where each possible genotype maps to a distinct phenotype), they must also revise their understanding of "dominance" and "recessiveness." Up to this point, most students tend to associate recessiveness either with (a) a phenotype; or (b) any genotype that contains a recessive allele (designated with the number 2); or (c) both. It is quite common for students to conclude that the phenotypes associated with (1,1) and (1,2) genotypes are both "recessive."2 However, this is inconsistent with their prior concept of recessiveness as it was developed in the simple dominance model. Thus, it is at this point in the unit that we emphasize the need for models to be consistent with other knowledge in a scientific discipline. In other words, geneticists must assess a new inheritance model, in part, based upon how well it fits within an existing family of related models such as meiosis (including cytological data) and molecular biology (which specifies the relationships between DNA and proteins as well as protein actions in cells). After explicit instruction about DNA transcription, translation, and protein function, students attempt to reconcile their codominance models with this new model of protein action in cells. In the case of codominance, this requires them to conceptualize recessiveness at the level of alleles and their relationships to one another rather than at the level of phenotypes or genotypes. In so doing, students construct meanings for dominance and recessiveness that are consistent across various inheritance models (i.e. simple dominance, codominance, multiple alleles, etc.) as well as models of meiosis and molecular biology.
For the final GCK inquiry, the students are organized into two research teams, each of which consists of four small research groups. Each team is assigned a population of virtual fruit flies and told to explain the inheritance of four traits within this population. The work is divided such that each research group studies two of the traits. Consequently, there is some overlap of trait assignments among the groups within a team. The teams hold research meetings periodically and a minimal structure for those meetings is imposed: Two groups present some data and tentative explanations of the data, one group moderates the meeting, and one group records the proceedings. The roles of individual groups alternate in successive meetings.
| The entire class then gathers for a final conference during which students create posters that summarize their research findings, take turns making formal presentations of their models, and critique their classmates' models. |
Each of the fly populations in this last problem contains traits that exhibit the following inheritance patterns: (1) Mendelian simple dominance; (2) codominance; (3) multiple alleles (specifically, three different alleles with varying dominant/codominant relationships between pairs of alleles); and (4) X-linkage. After about a week of data collection, model testing, and team meetings, each small research group is usually able to describe a model of inheritance for at least one of the traits in their population, and most groups can describe inheritance models for both of the traits on which they chose to focus. The entire class then gathers for a final conference during which students create posters that summarize their research findings, take turns making formal presentations of their models, and critique their classmates' models.
This high school biology curriculum is designed to give students opportunities to learn about genetic inquiry in part by providing them with realistic experiences doing inquiry in the discipline. Since a primary goal of practicing scientists is to construct explanatory models to account for natural phenomena, involving students in the construction of their own explanatory models provides a major emphasis in the classroom. The students work in groups structured like scientific communities to develop, revise, and defend explanatory models for inheritance phenomena. The overall instructional goals include helping students to understand mechanistic explanations for inheritance patterns in fruit flies and humans and to appreciate the degree to which scientists rely on empirical data as well as broader conceptual knowledge to assess explanatory models. The role that research about students' ideas has played in helping to achieve these instructional goals is the focus of the next section of this chapter.
Attending to Student Knowledge
The collaboration between high school science teachers and educational researchers that has recently come to be known as MUSE really began over a decade ago with a nine week genetics course in a biology curriculum for juniors and seniors. That original genetics course provided the setting for many studies about student learning, with a particular focus on how students used, revised, and judged scientific models. What we have learned about students in our own classrooms, as well as what others have shared in related studies (Kindfield, 1991 & 1994; Grosslight, et al., 1991; also see Allchin, 2000), has informed us as we-
- sequence concepts within the curriculum.
- design tasks for students & structure classroom instruction.
- design and implement formative and authentic assessments.
| Students begin their exploration of Mendelian inheritance in pea plants with a firm understanding of a basic meiotic model and continue to refer to this model as they examine increasingly complex inheritance patterns in humans. |
Curriculum. Students' difficulty in thinking about and discussing issues related to the consistency of particular genetics concepts with one another informed the sequence of elements within the genetics unit. In our early studies (as well as those in similar studies on problem solving in genetics-see Kindfield, 1994) students often did not examine their inheritance models to see whether they were consistent with meiosis. In fact, students proposed models where offspring received unequal amounts of genetic information from their two parents or had fewer alleles at a particular locus than did their parents (Cartier, 2000a). Because of their struggles and the fact that meiosis is central to any particular model of inheritance, we placed this model first in the revised curricular sequence. Students now begin their exploration of Mendelian inheritance with a firm understanding of a basic meiotic model and continue to refer to this model as they examine increasingly complex inheritance patterns. Another example of how the curricular sequence pushes students to examine ideas for consistency is the placement of instruction about molecular biology and protein function after students develop a codominant model of inheritance (described in detail in the previous section).
Instruction. Students' ideas about models. One early study of student learning in the genetics unit focused on identifying the criteria that students used when assessing their models for inheritance phenomena (Cartier, 2000a). The study was predicated on a commitment to developing the idea of consistency as a basis for model assessment with students early in the course. Thus, students read a mystery scenario involving a car accident and evaluated several explanations of the cause of the accident. Each explanation was problematic because it was either (1) inconsistent with some of the information they'd been given; (2) inconsistent with their prior knowledge about the world; or (3) unable to explain all of the information mentioned in the original scenario. Students discussed these explanations and their shortcomings and the teacher provided the language for talking about model assessment criteria: she instructed them to seek-
- explanatory power.
- predictive power (which was discussed but not applied to the accident scenario).
- internal consistency (among elements within the model).
- external consistency (between a model and one's prior knowledge or other models).
| Studies have shown that students view models in a "naïve realistic" manner (as objects or replicas of actual phenomena) rather than as conceptual structures that scientists use to explain data and ask questions about the natural world. Grosslight, 1991 |
Throughout the genetics unit, students were prompted to use these criteria to evaluate their own assessment of inheritance models. Despite the explicit emphasis on consistency as a criterion for model assessment, however, we found that very few students actually judged their models this way. Instead, students valued explanatory adequacy, visual simplicity, and "understandability" more strongly. A closer look at the work of students in this study showed that most of them-7 of the 8 interview participants-did not view models as conceptual structures, but as physical replicas, instructional tools, or visual representations. Other studies have also shown that students view models in this "naive realistic" manner rather than as conceptual structures that scientists use to explain data and ask questions about the natural world (Grosslight, 1991; also see Harrison and Treagust, 1998). Given this understanding of models, it was not surprising that our genetics students neither applied nor discussed the criterion for conceptual consistency within and among models. Moreover, the students in this study also participated in a "black box" activity (similar to the detergent box task described on page XX) where they proposed explanations for a data pattern associated with an imaginary box (described in Cartier, 2000a). When attempting to apply the model assessment criteria to their explanations for this pattern, several students treated "internal consistency" and "external consistency" literally: They evaluated the box's proposed internal components and the external phenomena (observations) separately. This confusion stemmed from students' prior understanding of concepts associated with vocabulary that we provided: Clearly "internal" and "external" were already meaningful to students and their prior knowledge took precedence over the new meanings with which we attempted to imbue these terms.
| Teachers present sample models that purport to explain the phenomena at hand and ask students to evaluate these models. Most commonly, students will describe the need for a model to-
Throughout the course, the instructor will return to these assessment criteria in each discussion about students' own inheritance models. |
Following this study, we altered the instruction in the genetics unit to take into consideration students' prior knowledge about models and particular vocabulary used to describe their attributes. Most importantly, we recognized the powerful prior ideas students brought with them about models as representational entities and explicitly addressed these ideas at the outset of the unit. In the genetics unit currently taught by members of our collaborative, teachers employ tasks early on that solicit students' ideas about scientific models (including a black box task and a reading/homework reflection about models) and explicitly define the term "model" as it will be used in the science unit. Additionally, teachers provide model assessment tasks to students and allow them to develop their own vocabulary for discussing shortcomings of particular explanations. Frequently, teachers will present sample models that purport to explain the phenomena at hand (most often the black box data patterns) and ask students to evaluate these models. Teachers create the models to have particular shortcomings in order to prompt discussion by students. Most commonly, students will describe the need for a model to explain all the data, predict new experimental outcomes, and be realistic (their term for conceptual consistency). Throughout the course, teachers will return to these assessment criteria in each discussion about students' own inheritance models. A subsequent study has shown that these instructional modifications (along with other curricular changes in the genetics unit) helped students to understand the conceptual nature of scientific models and learn how to evaluate them for consistency with other ideas (Cartier, 2000b).
| In early versions of the genetics course, students paid attention more to empirical issues than to consistency among models when assessing their new inheritance models. When the course was revised, we situated the study of inheritance patterns within the context of a single population of organisms. This emphasized the need for each inheritance model to be basically consistent with other models within genetics. |
Students' model revision heuristics.Several early studies of students' GCK work in our genetics unit revealed that students assessed their tentative models based primarily on empirical criteria rather than conceptual criteria (Cartier, 2000a & 2000b). Even when conceptual inconsistencies occurred between their proposed models and other models or biological knowledge, their primary focus was usually on how well a given model could explain the data at hand. Students frequently had difficulty recognizing specific inconsistencies between their models and meiosis or other biological knowledge such as the method of sex determination in humans. In some instances, students recognized that their models were inconsistent with other knowledge but were willing to overlook such inconsistencies when their models were judged to have adequate explanatory power. (For example, students sometimes proposed models to account for x-linkage inheritance patterns wherein a male organism simply could never be heterozygous. No explanation consistent with independent assortment in meiosis was given for this model.) Thus, students paid more attention to empirical than conceptual issues and tended to value empirical power over conceptual consistency in models where both criteria were brought to bear. In summary, the students' methods of model assessment differed from those of practicing geneticists
White & Frederiksen (1998) have described a middle school science curriculum designed to teach students about the nature of inquiry generally and the role that modeling plays in specific scientific inquiries. One aspect of the curriculum that had a measurable effect on its success was the emphasis on reflective assessment on the part of the students. Following modeling activities, students were asked to rate themselves and others in various categories, including "understanding the science", "understanding the processes of inquiry", being systematic", and "writing and communicating well" (p. 25). Involving the students in such an explicit evaluation task helped emphasize the importance of learning "about" inquiry and modeling in addition to learning how to "do" inquiry.
| Influenced by our research in the genetics unit, we built into the curriculum more tasks that require students to reflect, write, and discuss conceptual aspects of genetic modeling. These tasks include-
|
Our work developing tasks for students is also predicated on the importance of metacognitive reflection on their part. Influenced by our research in the genetics unit, we built into the curriculum more tasks that require students to reflect, write, and discuss conceptual aspects of genetic modeling. These tasks include journal writing, written self-assessments, homework assignments that require students to explain their reasoning (see Figure 3), and class presentations (both formal and informal). Most importantly, we created a complex problem, involving several different inheritance patterns, and asked the students to account for this new data while working in cooperative laboratory teams. As described above, the regular team interactions required students to be critical of their own thinking and that of others. Moreover, situating the study of these inheritance patterns within the context of a single population of organisms helped to emphasize the need for each inheritance model to be basically consistent with other models within genetics. We have found that in this new context, students are more successful at proposing explanatory models and have a better understanding of the conceptual nature of such scientific models (Cartier 2000a & 2000b).
Assessment Design. Related to the design of tasks is the design of good assessments; That is, we attempt to employ formative and authentic assessments throughout the unit. Because students struggle with conceptual problems in the genetics unit, we incorporate a number of assessments that require students to describe the relationship between models or ideas that they have learned (Figure 4A). Whenever possible, we design formal assessments as well as written classroom tasks that reflect the structure of students' work in the classroom. Our students spend a lot of their class time working in groups, pouring over data, and talking with one another about their ideas. Thus, assessments also require them to look at data, propose explanations, and describe the thinking that led to particular conclusions (Figure 4B).
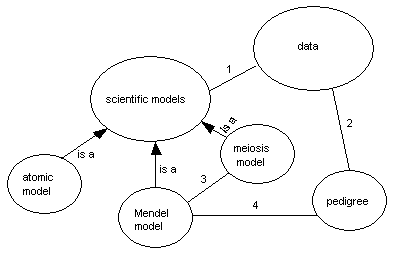
Figure 4aBelow is a concept map that represents the relationships between specific models, models in general, and data. Use the map to answer the questions below. a. Remember that a line in a concept map represents a relationship between two terms (concepts, ideas, etc.) in the map. Write a few sentences that describes the numbered relationships between the terms given. Be as specific as you can: use the appropriate vocabulary of genetics to make your point as clearly as possible. b. Draw a line (not necessarily a straight one) to separate the world of ideas from that of observations on this map. Please label both sides. Justify your placement of that line.
|
Figure 4bInheritance of PKU in the Samson Family
1. Use Mendel's simple dominance model to assign genotypes to the individuals in this pedigree. 2. Do the affected individuals in this pedigree show a dominant or recessive variation of the trait? Pick two family groups (a group is one set of parents and their offspring) and describe how those groups helped you make that decision. 3. Describe how you would convince another student, who had no knowledge of how PKU is inherited, that you understand the inheritance of this trait. As the student is not easily convinced, you must carefully show how the Mendel model can be used to support your idea. Figure 4B: Simple Dominance homework assignment. Students are asked to use Mendel's simple dominance model to explain a realistic data pattern. They are also asked to explicitly justify their reasongng similar to the way in which they argue in support of their ideas in regular classroom activities. |

Summary
The structure of the genetics class that we have described reflects important aspects of scientific practice: Students are engaged in an extended inquiry of inheritance through which they collect data, seek patterns and attempt to account for those patterns using explanatory models. The models proposed by students also bear a great deal of resemblance to those of practicing geneticists in that they specify allelic relationships and genotype to phenotype mappings for particular traits. In the next section, we will describe a course in evolutionary biology that provides opportunities for students to participate in realistic inquiry within another sub-discipline of biology. In the final section, we will discuss some important instructional and curricular elements that these two courses share and that are crucial to the learning success of students in these contexts.
Section 3: Developing Darwin's Model of Natural Selection in High School Evolution
| Developing a Darwinian Explanation Hillary and Jerome are sitting next to each other in their sixth hour science class waiting for the bell to ring.
The bell rings and their teacher announces that the class will start work on the last of three case studies designed to allow them to continue to develop and use the Darwinian model of natural selection. She tells the students that there are two parts to this third case. First, they will need to use their knowledge of the natural selection model to develop an explanation for the bright coloration of the male ring-necked pheasant. And secondly, they will have to write a research proposal that will then be considered by the rest of the students in a research grant competition.
As Mrs. J. passes out the eight pages of case materials she asks the students to get to work. Each group receives a file folder containing the task description and information about the natural history of the ring-necked pheasant. There are color pictures that show adult males, adult females, and young. Some of the pages contain information about predators, mating behavior and mating success. Hillary, Jerome and their third group member, Grace, begin to shuffle through the pages in the folder.
The three students spend the remainder of the period looking over and discussing various aspects of the case. By the middle of the period on Tuesday this group is just finalizing their explanation when Casey, a member of another group, asks if she can talk to them.
These two groups have very different explanations. Hillary's group is thinking that the males bright coloration distracts predators from the nest, while Casey's group has decided that the bright coloration confers an advantage on the males by helping them attract more mates. A lively discussion ensues.
|
The groups agree to disagree on their interpretation of this piece of data and continue to compare their explanations on other points.
| Because of the key role that natural selection plays in understanding evolution and biology more generally we devote a significant period of time to allow students to develop a deep understanding of the process. |
The students in the above vignette are using Darwin's model of natural selection and realistic data to create arguments about evolution in a population of organisms. In doing so they attend to and discuss ideas such as selective advantage and reproductive success that are crucial components of the model. Because of the key role that natural selection plays in understanding evolution and biology more generally we devote a significant period of time to allow students to develop a deep understanding of the process. Early in the course students have opportunities to learn about natural selection, but as the course progresses they are required to use their understanding to develop explanations (as illustrated in the vignette). This structure provides students with a setting in which to clarify their understanding of natural selection and it also gives them a context for learning by and about scientific inquiry. Section 3 will describe how the three design principles discussed earlier were influential in the development of this nine week course.
Disciplinary knowledge
| "There is probably no more original, more complex,
and bolder concept in the history of ideas than Darwin's mechanistic
explanation of adaptation." -Ernst Mayr (1982, p. 481) |
The choices we make when designing curricula are determined in part by examining the discipline under study. In the case of evolution it is clear that a solid understanding of natural selection provides a foundation upon which further knowledge can be built. But that foundation is hard won and takes time to develop because the concepts that comprise the natural selection model are difficult for students to understand and apply. Indeed, biologist Ernst Mayr has said that "there is probably no more original, more complex, and bolder concept in the history of ideas than Darwin's mechanistic explanation of adaptation" (1982, p. 481). Many courses or units in evolutionary biology at the high school level require far shorter periods of time than the nine weeks described here, and also include additional sophisticated concepts such as genetic drift and speciation. It seems clear to us, however, that with a large number of concepts covered in a short period of time the likelihood that students will develop a deep understanding of any of them is diminished-a survey of content is not sufficient to develop lasting understanding. Furthermore, we have expanded the notion of content to include opportunities for students to engage in and think explicitly about disciplinary inquiry. With these considerations in mind our teacher/researcher collaborative designed a course that requires students to not only learn about the natural selection model, but also to use it in ways that are analogous to those of evolutionary biologists.
| Our concept of a Darwinian explanation draws together the components of the natural selection model (shown in figure 5) with a narrative structure that demands attention to historical contingency. |
Central to our approach is a commitment to looking at authoritative accounts of scientific practice for guidance. Kitcher's (1994) concept of a Darwinian history assisted us in thinking about how students could use the natural selection model in authentic ways. He describes a Darwinian history as a "narrative which traces the successive modifications of a lineage of organisms from generation to generation in terms of various factors, most notably that of natural selection" (p. 20-21). The use of narrative explanation is a key way in which evolutionary biology can be distinguished from other scientific disciplines. "Narratives fix events along a temporal dimension, so that prior events are understood to have given rise to subsequent events and thereby explain them" (Richards, 1992). Thus, our concept of a Darwinian explanation draws together the components of the natural selection model (shown in figure 5) with a narrative structure that demands attention to historical contingency. Frequently textbook examples of explanations for particular traits take the form of what O'Hara (1988) terms "state-explanations"-that is, they explain the present day function of particular character states without reference to their history. In contrast, what we call a Darwinian explanation attempts to explain an event, or as O'Hara says, "how-possibly" a trait came into being. This type of explanation is summarized by Mayr (1997),
When a biologist tries to answer a question about a unique occurrence such as 'Why are there no hummingbirds in the Old World?' or 'Where did the species Homo sapiens originate?' he cannot rely on universal laws. The biologist has to study all the known facts relating to the particular problem, infer all sorts of consequences from the reconstructed constellation of factors, and then attempt to construct a scenario that would explain the observed facts in this particular case. In other words, he constructs a historical narrative (p. 64).
Although, our focus is limited in scope to the development and use of the natural selection model, these considerations of the form of explanation in the discipline played a key role in the design of the course described here.
| Providing opportunities for students to use the natural selection model to develop narrative explanations is a central goal of the course. |
Providing opportunities for students to use the natural selection model to develop narrative explanations that are consistent with the view described above is a central goal of the course. During the final weeks of the course, students are engaged in creating Darwinian explanations. In essence, students are asked to use the components of the natural selection model to make sense of realistic data that they have been given concerning an extant organism. Each scenario is presented to students as a case study and they are provided with materials that describe the natural history of the organism. Photographs, habitat and predator information, mating behavior and success, and phylogenetic data are examples of the types of information that may be included in a given case (complete case materials can be found on the MUSE website). Students then weave the information into a narrative that must take into account all of the components of a natural selection model and describe the change over time that may have occurred (see Figure 5 for a sample student Darwinian explanation). As students hone their abilities to develop and assess evolutionary arguments over three successive case studies, they are able to participate in realistic evolutionary inquiry.
Figure 5
Figure 5. Darwinian explanation written by a group of students at the end of the Monarch/Viceroy case |
| By the end of our course students are able to reason in sophisticated ways about evolutionary phenomena. |
By the end of our course students are able to reason in sophisticated ways about evolutionary phenomena. We believe that careful consideration of the discipline is an important part of realizing that goal, but also recognize that there are several other key areas that must be considered. In the sections that follow we describe how we structured the class to take seriously student ideas and how we developed a community in which students were required to make their own thinking explicit.
Student Knowledge
| Studies have confirmed what teachers already know-students have very tenacious misconceptions about the mechanism of evolution and its metaphysical assumptions. |
Anyone who has ever taught evolution can attest to the fact that students bring a wide range of conceptions and attitudes to the classroom. During the past two decades there have been several studies that document student ideas both before and after instruction (see for example Bishop and Anderson, 1990; Demastes, S., Good, R., Peebles, P. (1995, 1996) and Trowbridge & Cummins, 1992). These studies have confirmed what teachers already know-students have very tenacious misconceptions about the mechanism of evolution and its metaphysical assumptions. Our knowledge of the ideas students bring with them about both evolutionary biology and the nature of science has been important in our curriculum design efforts.
Curriculum: In the evolution course there are several specific ways in which we have considered student ideas when choosing and sequencing course content. For example, the course begins with an extended examination of argumentation in science and in evolution in particular. In early versions of the course we found that students were unable to engage in discussions about particular arguments because they did not have a framework with which they were all familiar. Our course has since been modified to provide opportunities for students to develop such a framework by participating in a cartoon sequencing activity (described in more detail below). Our decision to place this at the beginning of the course was a relatively easy one as we felt students needed to have the a framework for critiquing arguments for virtually every subsequent experience.
We have also found that students have difficulty in understanding the concept of variation that is a foundation of the natural selection model. These difficulties have been documented in the literature (Bishop & Anderson, 1990) and we have found them to be present in our own classrooms. Because of the experiences students have with variability in most genetics instruction-in which they usually examine traits with discrete variations-the concept of continuous variation can be a significant challenge for students. We have seen that an incomplete understanding of variation in populations promotes the student idea that adaptations are a result of a single dramatic mutation, or that selection is an all-or-none event operating on one of two to three possible phenotypes. Recognition of these problems has led to explicit instruction on variability in populations and perhaps more importantly the inclusion of opportunities for students to examine and characterize the variability present in real organisms before they begin using the concept in constructing Darwinian explanations.
One of the activities used for this purpose is a relatively simple one, but it provides a powerful visual representation that students can draw on later when thinking about variation in populations. Typically, students do not recognize the wide range of variation that is present even in familiar organisms. In order to give students experience thinking about and characterizing variation, we have them examine sunflower seeds. Their task is to count the stripes on a small sample, but even this simple direction is less than straightforward as the class then has to negotiate what counts as a stripe, whether to count one side or two, or if the edges should be included. Once they have come up with common criteria and have sorted their sample into small piles the teacher has them place their seeds into correspondingly numbered test tubes. The result, once the test tubes are lined up in a row, is a clear visual representation of a normal distribution. The subsequent discussion centers on ways to describe distributions using concepts like mean, median, and mode. This activity takes place prior to times when students will need to draw on their understanding of variation to construct explanations using the Darwinian model of natural selection.
| Careful thought has been given to how instruction can be carried out
to allow student ideas to be the focus in the classroom. The three
major sections of the course are each do this is their own way: • First, the students work with the teacher to develop a framework for examining arguments that focuses in part on prior knowledge. • Second, they examine historical ideas that correspond to common alternate conceptions students have. • Third, they work on case studies that require them to develop original explanations and defend them to their peers. |
| Students realize that they are all examining the same data but that they also each bring a lifetime of experience to the table. Together they establish that the process of making inferences is influenced by what they observe (the data) and by their own prior knowledge and beliefs. |
Instruction: Consideration of student knowledge is not simply a commitment we have when making curricular decisions; it also plays into the design of instructional tasks. An important feature of the evolution course is explicit discussion among students about their own ideas. Furthermore, we have designed instruction to address some of the common misconceptions that students bring to the study of evolution. We have given careful thought to how the actual instruction could be carried out to accomplish our goal of making student ideas a focus in the classroom.
The first activity in the evolution course is designed to develop a framework for examining arguments. This initial experience takes place in a non-science context in order to increase student comfort with the task. Students are given a set of cartoon frames that have been placed in random order. Their task is to work with their group to reconstruct a story using the information they can glean from the images. Students are enthusiastic about this task as they imagine how the images relate to one another and how they can all be tied together to create a story. The whole class then comes together to compare their 'stories' and discuss how they made decisions. The sequences presented by different groups usually vary quite a bit from group to group (see Figure 6 for examples of two sequences). This provides a context for a discussion of how inferences are made. The initial conversation centers on the observations they made of the images. However, it quickly becomes apparent that each person did not place the same importance on specific observations and that even though groups may have observed the same thing they may not have made the same decisions about the order of the cards. What ensues is a conversation about particular considerations that entered into their decision-making. Students realize that they are all examining the same images (the data) but that they also each bring a lifetime of experience with cartoons and stories to the table. Together they establish that the process of making inferences about the order of the cards is influenced by what they observe (the data) and by their own prior knowledge and beliefs. This notion is then generalized and students see that all inferences can be thought of as being based on these two things. They discuss how scientific arguments are usually a collection of several inferences that are all dependent on data and prior knowledge and beliefs. And finally they discuss the importance of questioning all three areas when critiquing an argument.
Figure 6Group One
"We think that in this first frame little red riding hood is telling the pigs that she is going to visit her sick grandmother. In the second scene, the pigs are telling the wolf about little red riding hood and her sick grandmother and showing him which way she went. In the next frame, the pigs see that the grandmother is tied up in the woods and they feel bad that they gave the wolf the information earlier." Group Two
"The pigs have discovered grandma tied up in the woods and they try to throw the wolf off the track by telling him that he must get away before the hunter comes. In the last frame, little red riding hood is thanking the pigs for saving the grandmother and they feel bashful." Figure 6. Two different groups of students interpreted the same images in different ways. There are thirteen images in the complete set that the students worked with for this activity. |



The cartoon activity described above provides the foundation for examining arguments that is used for the remainder of the course. It is important to note that an explicit part of that framework requires students to consider the prior knowledge and beliefs that they and others bring to bear on inferences made about data. In this way, the students are encouraged to continue to examine their own knowledge throughout the course.
| Students examine three historical models that account for species adaptation and diversity:
Once students have developed an understanding of the explanation that each author proposed and some familiarity with the observations on which it was based, they examine the prior knowledge and beliefs that each author may have held. |
The second major section of the evolution course engages students in examining three historical models that account for species adaptation and diversity. The students must draw on the framework established during the cartoon activity in order to accomplish this comparison. This means that as they examine each argument they also identify the major inferences made and the data and prior knowledge and beliefs that formed the basis of the inferences. The three models are: (a) William Paley's model of intelligent design that asserts that all organisms were made perfectly for their function by an intelligent creator, (b) Jean Baptiste de Lamarck's model of acquired characteristics that is based on a view that adaptations can result from the use or disuse of body parts and that changes accumulated during an organism's lifetime will be passed on to offspring, and (c) Charles Darwin's model of natural selection. The models of Paley and Lamarck were chosen because they each represent some of the common ideas that students bring with them to the classroom. Specifically, it is clear that many students attribute evolutionary change to the needs of an organism and believe that extended exposure to particular environments will result in lasting morphological change. Many students are also confused about the role of supernatural forces in evolution. Darwin's model is included in the analysis so that students can see how the underlying assumptions of his model compare to those of Paley and Lamarck.
| Examining Scientists' Beliefs
and Assumptions
Claire and Casey are working with Hillary in a group during class. They are trying to analyze and understand an excerpt of original writing by Lamarck. Hillary is looking over the discussion questions: Hillary: It seems like Lamarck did think that species changed over time, so I can see that as an underlying assumption of his, but I'm having a hard time figuring out how he thought that happened. Casey: I agree, he is definitely different from Paley who didn't think things had changed at all. Claire: But how did the change happen? It seems like Lamarck puts it on the organisms themselves, that they try to change. Hillary: I'm not sure what you mean. Claire: Well, he talks a lot about the usefulness of particular traits for an animal and about repeated use of a body part causing a change. |
| Even if a model can account for diverse phenomena on its own terms, it is still necessary to examine and critique the underlying assumptions. |
In order for students to compare the prior knowledge and beliefs of the authors, however, they first have to become familiar with the models. To do this, each model is taken in turn and students are discouraged from making comparisons until each model has been fully explored. All three models are presented in the same way. Students read edited selections of the author's original writing, answer questions about the reading and participate in a class discussion in which the proposed explanation for species diversity and adaptation is clarified and elaborated. Students are also given an opportunity to explore the natural phenomena or data that served as an inspiration for each author: they examine fossils as discussed by Lamarck, dissect an eye to examine the structure/function relationships that so fascinated Paley, and are visited by a pigeon breeder who brings several of the pigeon varieties that Darwin described in Origin of Species. Once students have developed an understanding of the explanation that each author proposed and some familiarity with the observations on which it was based, they examine the readings again in order to identify the prior knowledge and beliefs that each author may have held.
Following this discussion, the students compare the three models. First, students assess the explanatory power of the models, using each model to explain phenomena other than those described in the original writings. For example, they attempt to use Paley's model to explain the presence of fossils and Lamarck's model to explain the structure of the eye. Sometimes the model can easily account for new phenomena-Lamarck's model of use inheritance is easily adapted to pigeon varieties. In other instances, the students recognize the limitations of the model-Paley's model cannot easily account for the presence of fossils or extinct organisms. They then compare the underlying assumptions or beliefs of the authors. Even if a model can account for diverse phenomena on its own terms, it is still necessary to examine and critique the underlying assumptions. Many students question the necessity of the supernatural force underlying Paley's model, and still more students find the role of need to be a questionable assumption in Lamarck's model.
These explicit discussions of some of the major views that students bring to the study of evolution lay the groundwork for future use and extension of Darwin's model. The comparison of the assumptions of the three models enables students to distinguish between those beliefs that underlie the model of natural selection and those that do not. However, unlike some classroom contexts in which it is the students' ideas that are laid bare and examined for inconsistencies, here we have developed a situation where student ideas are represented by the models of Paley and Lamarck. In this way we have found that students are willing to attend to the differences between ideas rather than spending their time and energy being defensive. Because they do not feel that their own ideas are being criticized, the discussions are fruitful.
| Instruction emphasizes that assumptions of supernatural influence and individual need are not a part of the Darwinian model. Thus, when students use the natural selection model to explain natural phenomena, they often challenge each other when need-based or Lamarckian language is used. |
These two activities foster a classroom community that operates from a common set of metaphysical commitments. For our purposes, the most important of these is that Darwin proposed a naturalistic mechanism of species change that acts on variation among individuals within a species and that assumptions of supernatural influence and individual need are not a part of his model. Keeping this distinction in mind while using the natural selection model later in the course enables students to avoid some common misconceptions, or at least makes identification of those misconceptions more straightforward. For example, when students use the natural selection model to explain natural phenomena like the bright coloration of the monarch butterfly, they often challenge each other when need-based or Lamarckian language is used.
This type of self or peer monitoring becomes particularly obvious during the last section of the course when students are engaged in using the natural selection model to explain natural phenomena presented as case studies. For example, the second case requires students to provide a Darwinian explanation for the similarity in color between the monarch and viceroy butterflies. Frequently students will say things like, "the viceroy needs to look like the monarch so that the birds won't eat it." When statements such as these are made, other students will often challenge the speaker to use Darwinian rather than Lamarckian language. The work on the cases allows students to practice using the Darwinian model in appropriate ways and the interactive nature of all of the work in class affords them opportunities to think explicitly about and defend their own ideas.
Assessment Students' consideration of their own ideas has been incorporated in the assessment tasks in the evolution course. On several occasions and in different ways students examine their own ideas and explicitly discuss how those ideas have changed. For example, one of the questions on the final exam requires them to read and critique a Darwinian explanation they created on the first day of class. We have found this to be one of the most powerful moments for many students as they recognize how much their own ideas have changed. For example, many students are critical of the need-based language that was present in their original explanation or they find that they described evolutionary change as having happened at the individual rather than the population level.
Figure 7Original answer The saddleback carapace came into being due to the need of migrating tortoises to adjust to a new environment. On Albermarle Island the domed shaped carapaces served well for shedding rain and eating ground vegetation. However, when the tortoises began to migrate to smaller, drier island with less ground vegetation, they had to adapt in order to survive. The majority of the food was now higher up and the domed shell served as a hindrance. Over time, the saddleback carapace developed to allow the neck to extend further, thereby allowing the tortoises to reach the fleshy green parts of the prickly pear cactus. This evolutionary process created a new species of giant tortoise that could live successfully in a new environment. Critique on final exam In my original answer, I used an almost exclusive Lamarckian definition of evolution. In my introductory statement I stated that the saddleback carapace came into being due to the need of the tortoise to fit its environment. I needed to acknowledge the existence of variation within the tortoise population of the shape of the shell. My original explanation makes the evolutionary process sound like a physical change taking place during the life of the tortoise and then being passed on to the offspring. I now know that variations that are advantageous give animals a better chance of survival (survival of the fittest!) and allow them a better chance of passing on their advantageous trait to their offsping. In my original explanation I also touched on ideas of use and disuse to explain how the saddleback carapace came to be, this is a Lamarkian model of evolution which is incorrect. I did explain how the saddleback carapace was an advantage because it allowed the tortoise to eat higher vegetation. Since I didn't understand evolution through the generations, I wasn't able to describe how the species changed over time. Overall, I would say I had a basic but flawed understanding of evolution but I lacked the tools to explain evolution from a scientific and Darwinian perspective, until now. Figure 7. On the first day of class, students were asked to explain how the carapace shape of Galapagos tortoises may have changed from the dome-shape to the saddleback shape. As part of the final exam, students were asked to critique the explanation they gave on the first day. Above is the original explanation and the critique offered by one student. |
| We have found that the most effective assessment tasks are those that most closely resemble the ways in which students learned the science. |
| One way to assess student understanding is to provide them with novel data and ask them to describe their reasoning about that data using the natural selection model. |
Additionally, we have found that the most effective assessment tasks are those that most closely resemble the ways in which students learned the material. In other words, we have found that students are more successful and the data we collect on what they know is more complete and accurate when they are engaged in tasks that are analogous to the instruction they experienced. For example, during instruction students are required to use the natural selection model to develop Darwinian explanations that account for rich data sets. To then ask them about data or the components of natural selection in a multiple choice format that would only require them to draw on bits and pieces of knowledge for any one question seems incomplete at best. Instead, we provide them with novel data and ask them to describe their reasoning about that data using the natural selection model-a task that is analogous to what they have been doing in class. An instance of this type of assessment on the final exam asks students to write a Darwinian explanation for polar bear fur color using information about ancestral populations. In this way, during assessment, we draw on student ideas and skills as they were developed in class rather than asking students to simply recall bits of information in contrived testing situations.
Section 4: Putting It All Together - Promoting Reflective Scientific Practice in Classrooms
| The model-based inquiry curricula are designed to facilitate reflective student thinking through-
|
| Typically during the game of school students "win" when they present the correct answer. The process by which one determines the answer is irrelevant or, at best, undervalued. |
We have found that much of what students learn in model-based inquiry genetics and evolutionary biology units depends upon their active and thoughtful participation in the classroom community(Cartier, 2000a & 2000b; Passmore & Stewart, 2000; Passmore & Stewart, in press). In order to learn about the process of modeling and about discipline-specific patterns of argumentation as they participate in it, students must be critically aware of the elements that influence their own knowledge generation or justification. The MUSE curricula are designed to facilitate this type of student thinking through explicit discussion of student expectations for engaging in argumentation, the design of student tasks, and the use of various tools for interacting with and representing abstract concepts.
Argumentation
High school students have had more than nine years of practice playing the "game of school." Most have become quite adept at memorizing and reiterating information, seeking answers to questions or problems, and moving quickly from one topic to another. Typically during the game of school, students "win" when they present the correct answer. The process by which one determines the answer is irrelevant or, at best, undervalued. The students described here are quite typical in this regard: They enter the genetics and evolution classes anticipating that they will be called upon to provide answers and are prepared to do so. In fact, seeking an end product is so ingrained that even when we design tasks that involve multiple iterations of modeling and testing ideas, such as within the genetics course, students frequently reduce the work to seeking algorithms that have predictive power rather than engage in the much harder task of evaluating models based upon their conceptual consistency within a family of related ideas (see Cartier, 2000a; and discussion in Section 2 pg. X, "Students' Model Revision Heuristics").
| Expectations for the Evolution Class On the first day of instruction in the evolution course, students are made aware that the expectations may be different from those they have experienced before. Students are given a summary of the goals of the course and the teacher discusses this with them. This summary states that, "To help you understand how biologists reason to address questions from an evolutionary perspective you will have the opportunity to work on a variety of realistic problems…you will be responsible for studying these problems in small groups, and presenting your solutions to your classmates…this class is all about making sense out of what we know and this requires people to continually question each others' understanding in constructive ways…it will require you to justify statements that you make and question the statements of others" Through this handout and subsequent guidance of the teacher, students realize that they will have to seriously consider their ideas and those of others. |
After studying how people solved problems in a variety of situations, Klayman & Ha (1987) noted the frequent use of what they called a "positive test strategy." That is, solvers would propose a model (or solution) and test it by attempting to apply it to the situation most likely to fit the model in the first place. If the idea had explanatory or predictive power, the solver remained satisfied with it; if not, the solver would quickly test another idea. The positive test strategy was frequently applied by students in early versions of the genetics course (Cartier, 2000a). This method of problem solving does not map well onto scientific practice in most cases, however: It is the absence of disproving evidence, and not the presence of confirming evidence, that is more commonly persuasive to scientists. Moreover, testing a model in limited situations where one expects data-model match would be considered "confirmation bias" within scientific communities. Nevertheless, Klayman and Ha point out that this positive test strategy is often quite useful in real life situations and they note that "in the absence of specific information that defines some tests as more relevant than others" it is frequently used.
Given our students' facility with the game of school and the general tendency to apply less scientific model-testing strategies when problem solving, we are confronted with the need to create tasks that will not only afford the opportunity for reflection, but actually require students to think more deeply about the ways in which they came to understand science concepts as well as what is involved in scientific argumentation in the courses described here. We want students to realize that "claims . . . are open to challenge and progress is made through dispute, conflict, and . . . change" in science (Driver, et al., 2000). Thus, we explicitly discuss with our students the expectations for their participation in the course. Teachers state that the students' task is not simply to produce an "answer" (a model in genetics, or a Darwinian explanation in evolutionary biology), but to be able to defend and critique ideas according to the norms of a particular scientific discipline. In other words, we ask the students to abandon the "game of school" and begin to play the "game of science."
Examination of ideas requires more than simply providing space for reflection to occur, it also involves working with students to develop systematic ways to critique their own ideas and those of others. Thus, we begin each course with an activity whose focus is the introduction of discipline-specific ways of generating and critiquing knowledge claims. The activities are frequently described as "science free" because there is no expectation that students will come to understand any particular scientific concepts upon completion. Rather, they will have learned about the process of constructing and evaluating arguments in genetics or evolutionary biology. Specific criteria for weighing scientific explanations are returned to throughout each course as students engage in extended inquiries within these biological disciplines.
Our discussion of student expectations for engaging in argumentation will be illustrated with examples drawn from the genetics and the evolution classes.
| The black box activity creates many opportunities to introduce and reinforce foundational ideas about the nature of scientific inquiry and how one judges ideas in genetics. |
Genetics Course: The Black Box Activity. The first activity of the genetics unit, the "black box activity", is designed to introduce students to appropriate ways of discussing and assessing ideas This activity shares some important characteristics with the cartoon activity (used to begin the evolution course), in that it provides a context within which to discuss criteria for judging scientific ideas. However, genetic arguments differ from those in evolution in a number of ways, including the emphasis on demonstrative evidence in genetics that is absent (for the most part) from arguments in evolutionary biology (see Rudolph and Stewart, 1998).
During the first few days of the genetics course, the teacher presents the students with a "black box"-either an actual box or a diagram and description of a hypothetical box-and demonstrates or describes the phenomenon associated with it. For example, one "box" is a cardboard detergent container that dispenses a set amount of detergent each time it is tipped; another is a large wooden box with a funnel on top and an outlet tube at the bottom that dispenses water in varying amounts. Once the students have had an opportunity to establish the data pattern associated with the particular box in question, the teacher explains that the students' task is to determine what mechanisms might give rise to this observable pattern. During this activity, (which can take anywhere from 3 to 11 class periods, depending upon the black box that is used and the extent to which students can collect their own data), the students work in small teams (see Figure 8 for an example of another "black box". At the conclusion of the task, each team creates a poster representing their explanation for the box mechanism and presents it to the class. Classmates offer criticism and seek clarification during these presentations.
Figure 8
Figure 8: One "black box" used in the MUSE science curriculum and typical data patterns associated with the box. |

This activity creates many opportunities to introduce and reinforce foundational ideas about the nature of scientific inquiry (modeling) and how one judges ideas in genetics. As the class shares early ideas, the teacher leads discussion about the criteria they are using to decide whether and how to modify these initial explanations. Together, the class establishes that an explanation-which at this point is renamed a "scientific model"-must be able to explain the data at hand, accurately predict the results of future experiments, and be consistent with prior knowledge (or be "realistic"). Through discussion and a short reading about scientific inquiry and model assessment, the teacher helps students to connect their own work on the black boxes with that of scientists attempting to understand how the natural world works. This framework for thinking about scientific inquiry and determining the validity of knowledge claims is returned to repeatedly throughout the genetics unit.
| The cartoon activity introduces students to evolution-specific norms of argumentation. |
Evolution Course: The Cartoon Activity. The first activity of the evolution course is the cartoon activity (described in Section 3). It is used to exemplify how to critically examine arguments and to think explicitly about one's own ideas. It also serves to establish general norms of interaction that encourage student discussion and debate. During student presentations of the cartoon sequences, the teacher points out examples of fruitful questioning and encourages the students to think about what it means to foster a community where communication about important ideas is expected. These norms are then reinforced during the remainder of the course.
In addition to introducing general norms for classrooms where scientific argumentation is central, the cartoon activity serves to orient students to a framework for critiquing arguments in evolution. At one level this framework is common to all science disciplines. In this capacity the emphasis is on the importance of being explicit about how prior knowledge and beliefs influence the inferences drawn from particular data. At this general level the activity is connected to the common MUSE framework of models and modeling as the teacher connects the ideas they have discussed concerning inferences to those of models. The teacher does this by explaining that a scientific model is an idea that accounts for some set of phenomena and that models are based on several inferences. Students then do a reading about models and as a class they discuss the ways in which models can be assessed. Through examples in the reading and from their own experience the group settles on criteria for judging models: Explanatory power, and consistency with other knowledge. Note though that unlike the genetics course, there is no mention of predictive adequacy here as a major assessment criterion. This is because explanation is much more central than prediction in the evolution course. This is one example of the assertion that we have made before-disciplines do rely on differing methods for making and evaluating claims. The demonstrative inference that is common in the genetics course gives way to a greater reliance on non-demonstrative inference in the evolution course. This occurs as students create functional Darwinian explanations. Such explanations, with their characteristic narrative structure, are developed to make sense out of disparate date (structural and behavioral characteristics of organisms, patterns in their molecular biology, patterns of distribution in both time and geography and so on) that is most characteristic of evolutionary argumentation.
A second evolution-specific function served by the cartoon activity is to introduce students to one of the more important undertakings of evolutionary biologists-the reconstruction of past events (the development of a trait, i.e. the vertebrate eye; or the speciation events that led to the "bush of life"). Such historical reconstructions do not have close analogues in genetic inquiry.
Tasks
| Discourse is anchored in norms of argumentation that reflect scientific practice to the highest degree possible. |
In addition to establishing expectations for class participation and a basic framework for knowledge assessment, these biology curricula promote metacognitive reflection on the part of students by incorporating tasks that require discourse (formal and informal) at all stages of student work. While working in groups and presenting results to the class as a whole, students are required to share their ideas even when those ideas might not be fully formed. Moreover, recall that the context for idea sharing is one in which discipline-specific criteria for assessment of ideas has been established. Thus, discourse is anchored in norms of argumentation that reflect scientific practice to the highest degree possible. An example from the evolution course will serve to illustrate the design of tasks in a model-based inquiry curricula..
| The evolution class is designed to encourage collaboration and sharing among groups. |
Developing Darwinian Histories. One of the major tasks students engage in during the evolution course is the production of Darwinian explanations for realistic situations. To this end, the second half of the course is dedicated to three different case studies. In the first of these students develop a Darwinian explanation for differences in seed coat characteristics among populations of a hypothetical plant species. The second involves explaining the bright, and similar, coloration of Monarch and Viceroy butterflies, and the final case requires that students develop an explanation for how the sexual dimorphism exhibited by ring-necked pheasants might have arisen. As is typical in the classrooms described here, students share their explanations at the culmination of their work. Yet, of equal importance is the sharing they do as they develop their explanations.
| The students normally propose more than a single explanation for a phenomena, thus recognizing that it is important to consider 'multiple interpretations. |
During each case study the work time is structured so that a group will consult with at least one other group as they develop their explanations. This task organization reinforces the nature of argumentation in evolutionary biology as it includes the expectation that students will attend to the central feature of any Darwinian explanation-that there be an historical component to it. But, it is not enough to just have a history. In tracing the possible historical development of a trait students must weave a complex story that draws on available data as well as their understanding of an array of biological models (e.g. genetic ones) to discuss the role of heritable variation, super fecundity, competition, and agents of selection. Within their research groups, meetings between research groups, and whole-class discussions students question one another using a variety of sophisticated stances. These include assuring that there is consistency between data, the natural selection model, and claims; that the history of the shift in a trait is feasible (i.e., consistent with genetics); and that the proposed selection agent could have brought about the change in the trait between times 1 and 2. The students question one another to insure that their explanation is both internally and externally consistent. In doing so they normally propose more than a single explanation, thus recognizing that, in evolution at least, it is important to consider "multiple interpretations." As they examine the competing Darwinian explanations for the same phenomena they invoke evolution-specific argument-analysis norms-that the explanation of the history of a trait has to be consistent with the natural selection model.
Public Sharing of Ideas. The culminating activities for each of the three cases require public sharing of ideas in a forum where the expectation is that the presenting groups and audience members thoughtfully consider the ideas before them. Each case has a different type of final presentation. The first case ends with a poster session, the second with a round table discussion, and the last with an oral presentation.
| Students construct a Darwinian explanation for sexual dimorphism in ring-necked pheasants and, in addition, they also develop a research proposal to shed light on their explanation. |
One particularly powerful experience students have during the evolution course occurs during the final case. For the first two case studies, students use their understanding of the Darwinian model to account for the changes that may have occurred in particular populations and to explicitly tie data from the case materials to their claims. For the final case study they must construct a Darwinian explanation for the sexual dimorphism observed between male and female ring-necked pheasants and, in addition, they must also produce a research proposal to shed light on their explanation. Typically, students choose to focus their research proposal on a single aspect of their explanation. This activity requires that they think carefully about the components of their explanations and the confidence they place in each of those components. So in this instance they are not evaluating the entire explanation as a single entity, but rather are considering each part in relation to the others. Once they have decided on a research proposal they must then determine how their proposed research would strengthen their argument. Being able to examine an argument in part and as a whole is an important skill that this task helps develop. This case also stimulates interesting conversations among groups. Here the basic explanations may differ and therefore to understand the research proposals it is necessary for students to attend to the details of other groups' explanations. The non-presenting groups act as a proposal review panel and interact with the presenting groups in an attempt to understand the proposal. Once all groups have presented the students discuss the merits and shortcomings of each proposal and then individually decide which proposal to fund.
Tools
| In order for students to communicate with one another, they need to establish some meaningful language and vocabulary, useful notational systems to convey scientific concepts, and agreed-upon conventions to represent ideas. |
Finally, in order for students to communicate with one another, they need to establish some meaningful language and vocabulary, useful notational systems to convey scientific concepts, and agreed-upon conventions to represent ideas. The importance of attending to language, notation, and representations stems from the realization that every scientific community, in order to function, has created or co-opted exemplars of these three tool categories. Without them the ability to frame inquiries, analyze data, or make and critique claims would be compromised or rendered impossible. We will illustrate the function of tools within a modeling classroom with an example taken from the genetics instruction.
There is a literature that makes the case for the use of tools (language, notation, representation) to scaffold students' understanding and abilities to engage in discipline-specific argumentation. In fact, Lehrer, Schauble, Carpenter, and Penner (2000) who study the role of tools in facilitating science and mathematics learning in young students, have gone so far as to claim that, "
We tracked the evolution and interdependence of forms of inscription [these encompass notational systems and a variety of representations] and found that classroom episodes are characterized by a "cascade" of inscription in which the natural world is understood through a history of more powerful inscriptions. As children's inscriptions evolved, so too did their understanding of the world, and transformations in understanding were accompanied by corresponding transformations in their systems of inscription (pg. 2).
| For understanding to develop it is important to attend to the development of instructional tasks and activities that introduce students to powerful domain-specific tools that promote thinking, reasoning, and argumentation. |
Palincsar and Magnusson (199x; see also this volume pg. x)have created a scientists notebook as a part of their innovative science instruction for elementary school children. The notebook incorporates each of the three tools that we will describe below as a way to initiate young students into the "game of science." Similarly, Kindfield (1991; 1994) found that individuals who were more advanced in their understanding of genetics
"… systematically used their fine-tuned diagrams [a representation] as 'tools' to think with while reasoning." She goes on to say that "… more advanced pictorial skills [representations] and conceptual knowledge coevolve or mutually influence one another in the development of understanding of a subcellular biological process" (pg. 1).
Collectively these authors make the case that for understanding to develop it is important to attend to the development of instructional tasks and activities that introduce students to powerful domain-specific tools that promote thinking, reasoning, and argumentation. We have, in our approach to developing courses and units with a model-based inquiry approach, adopted a similar stance as a way to insure that science classrooms are places of reflective engagement for students.
Our intention for students is that they become members of a classroom community of inquirers and that they begin to understand something about what it means to do science. To accomplish this it has proven necessary to pay careful attention to the development of classroom-appropriate tools.
Language. Rather than provide an enormous glossary of terms as most text books do, we attempt to limit the number of technical terms to only those necessary for students to share their ideas. For example, while most text books include the terms "heterozygote" and "homozygote" in materials about inheritance, we introduce only the term "allele" and students themselves invent ways to talk about "people with two different alleles" and "people with two of the same kind of allele." This may seem less efficient than making use of common scientific jargon, but we have found that, rather than being helped by a lot of precise vocabulary, students are frequently confused by it. Similarly, we establish some basic vocabulary for talking about model assessment: "explain," "predict," and "realistic" are terms that students use to describe their assessment process during the Black Box activity and ones that are repeatedly used throughout the unit.
Notation. Perhaps more important than vocabulary in the genetics unit is the notational system that students use to describe alleles, their movement through meiosis, and the phenotypes that result from particular combinations of alleles. In the genetics community, the convention for representing alleles and genotypes varies depending upon the organism that one studies. To avoid potential confusion and to emphasize the universality of inheritance mechanisms throughout the biological world, we have established a "generic" notational system to represent genotypes. In our system, alleles are represented by numbers, with lower numbers usually assigned to alleles that are dominant. For example, in the simple dominance model, (see Figure 6), the two different alleles in the population are represented as 1 and 2, with allele 1 being dominant to allele 2. Making use of this notational system enables students to readily recognize inheritance patterns regardless of the particular trait involved: that is, students understand that a person possessing sickle cell trait has the genotype (1,2), as does a female who carries the colorblindness gene. More traditional ways of noting these genotypes would be (Ha, Hs) and (Cwt, C*), respectively. Clearly, it is easier to note the conceptual similarities (i.e. that each of these individuals possesses genetic information for two different alleles for the traits in question) when a universal notational system is employed. Moreover, the fact that both of these individuals have (1,2) genotypes at these loci begs the question of why a (1,2) genotype at the hemoglobin locus results in an observable phenotype at the cytological level but a (1,2) genotype at the colorblindness locus does not. Such comparisons pave the way for discussions about gene products (proteins) and their various functions in cells and organisms, as well as a revisitation of the concepts of "dominance" and "recessiveness."
| Students need concrete representational tools in order to be able to discuss abstract entities. |
Representation. Representational tools have proven quite valuable in the genetics unit, as students are reasoning and communicating about abstract entities. Consequently, we employ a number of such tools throughout the unit. While students are studying meiosis, gamete formation, and genotype to phenotype mappings early in the unit, they manipulate laminated, color-coded, paper pea plant "chromosomes" and simulate the processes of meiosis (of four chromosome pairs) and fertilization. Furthermore, they use pipe cleaners and laminated paper leaves, pods, and flowers to construct the plants the result from these processes. This activity provides students with concrete representational tools to discuss abstract entities (like alleles) and helps them to connect the processes of meiosis and fertilization with resulting phenotypes in whole organisms.
Another representational tool we utilize throughout the genetics unit is the GCK software. This program enables students to simulate the process of reproduction and observe the resultant "organisms" at the phenotypic level. Thus, unlike work with actual organisms, the students can perform matings and observe results almost instantly, allowing them to test whether proposed inheritance models can be used to accurately predict.
SUMMARY
| For students to develop understanding in any scientific discipline requires that teachers and curriculum developers attend to a set of complex interrelated components. |
| Scientific literacy involves understanding central ideas in disciplines and how those ideas, and the claims that support them, have been generated and justified. |
For students to develop understanding in any scientific discipline teachers and curriculum developers must attend to a set of complex and interrelated components. These include the nature of practice in particular scientific disciplines, student prior knowledge, and the establishment of a collaborative environment for inquiry. These design components allow educators to create curricula and instructional materials that help students to learn about science as and by inquiry.
The students in the biology classrooms we've described have developed sophisticated understanding of some of the most central explanatory frameworks in genetics and in evolutionary biology. In addition, they have, unlike many high school students, shown great maturity in their abilities to reason about realistic biological data and phenomena with these central models. Moreover, they have accomplished this in classrooms that are structured along the lines of scientific communities. For this to happen there has been a concerted collaboration between high school teachers and their students, university science educators, and university biologists. That MUSE combined these collaborative elements with a research program on student learning and reasoning has been essential. With the knowledge gained from this collaboration we feel that it is possible to help others to realize the expectations for improving science education that are laid out in reform documents such as the National Science Education Standards. In particular these documents have called for curricular reforms that allow students to be "engaged in inquiry" that involves "combin[ing] processes and scientific knowledge as they use scientific reasoning and critical thinking to develop their understanding of science." (p. 105). Recommendations for improved teaching of science are solidly rooted in a commitment to teaching both through and about inquiry. Furthermore, the Standards don't simply suggest that science teachers incorporate inquiry in classrooms; rather, they demand that teachers embrace inquiry from a pedagogical standpoint: "Teachers of science-
- plan an inquiry based science program for their students.
- focus and support inquiries while interacting with students.
- create a setting for student work that is flexible and supportive of science inquiry.
- model and emphasize the skills, attitudes, and values of scientific inquiry." ( NRC, pp. 27-52)
It is just these opportunities that have been described in this chapter.
References
Allchin, D. (2000). Mending mendelism. The American Biology Teacher, 62 (9), 633-639.
American Association for the Advancement of Science. (1993). Benchmarks for science literacy: Project 2061. New York: Oxford University Press.
Bishop, B. A., & Anderson, C. W. (1990). Student conceptions of natural selection and its role in evolution. Journal of Research in Science Teaching, 27 (5), 415-427.
Calley J., & Jungck, J.R. (1997) genetics construction kit (The BioQUEST Library IV, version 1.1B3) [Computer software]. The ePress Project, College Park.
Cartier, J. L. (2000a). Assessment of explanatory models in genetics: Insights into students' conceptions of scientific models. Research report 98-1 for the National Center for Improving Student Learning & Achievement in Mathematics & Science. (http://ncisla.wceruw.org/publications/reports/RR98-1.PDF)
Cartier, J. L. (2000b). Using a modeling approach to explore scientific epistemology with high school biology students. Research report 99-1 for the National Center for Improving Student Learning & Achievement in Mathematics & Science. (http://ncisla.wceruw.org/publications/reports/RR99-1.PDF)
Cartier, J., & Stewart, J. (2000). Teaching the nature of inquiry: further development in a high school genetics curriculum. Science and Education, 9, 247-267.
Cartier, J., Rudolph, J., & Stewart, J, (2001). The nature and structure of scientific models. Working paper available through the National Center for Improving Student Learning & Achievement in Mathematics & Science. (http://ncisla.wceruw.org/publications/reports/Models.pdf)
Clough, E. E., & Wood-Robinson, C. (1985a). How secondary students interpret instances of biological adaptation. Journal of Biological Education, 19 (4), 304-310.
Clough, E.E. & Wood-Robinson, C. (1985b). Children's understanding of inheritance. Journal of Biological Education, 19(4), 304-310.
Darden, L. (1991). Theory change in science: Strategies from mendelian genetics. Oxford University Press, New York.
Demastes, S. S., Trowbridge, J. E., & Cummins, C. L. (1992). Resource paper on evolution education research. In R. G. Good, J. E. Trowbridge, S. S. Demastes, J. H. Wandersee, M. S.
Demastes, S., Good, R., Peebles, P. (1995), Students' conceptual ecologies and the process of conceptual change in evolution. Science Education, 79 (6), 637-666.
Demastes, S., Good, R., Peebles, P. (1996). Patterns of conceptual change in evolution. Journal of Research in Science Teaching, 33(4), 407-431.
Driver, R., Newton, P., & Osborne, J. (2000). Establishing the norms of scientific argumentation. Science Education, 84, 287-312.
Grosslight, L., Unger, C., Jay, E., & Smith, C. L. (1991). Understanding models and their use in science: Conceptions of middle and high school students and experts. Journal of Research in Science Teaching, 28, 799-822.
Hafner, & C. L. Cummins (Eds.), (1992) Proceedings of the 1992 evolution education research conference, (pp. 42-77). Baton Rouge, LA: Louisiana State University.
Hafner, R., & Culp, S. (1991). Elaborating the structures of a science discipline to improve problem-solving instruction: An account of classical genetics' theory structure, function, and development. Science and Education, 5, 331-355.
Harrison, A.G., & Treagust, D.F. (1998). Modelling in science lessons: Are there better ways to learn with models? School Science and Mathematics, 98 (8), 420-429.
Jensen, M. S., & Finley, F. (1995). Teaching evolution using historical arguments in a conceptual change strategy. Science Education, 79 (2), 146-166.
Jensen, M. S., & Finley, F. (1996). Changes in student's understanding of evolution resulting from different curricular and instructional strategies. Journal of Research in Science Teaching, 33 (8) 879-900.
Kitcher, P. (1993). The advancement of science: Science without legend, objectivity without illusions. New York: Oxford University Press
Kitcher, P. (1984). 1953 and all that. A tale of two sciences. The Philosophical Review, 93, 335-373.
Kindfield, A. C. H. (1991). Confusing chromosome number and structure: A common student error. Journal of Biological Education, 25(3), 193-200.
Kindfield, A. C. H. (1994). Understanding a basic biological process: Expert and novice models of meiosis. Science Education, 78(3), 255-283.
Klayman, J. & Ha, Y. (1987). Confirmation, disconfirmation, and information in hypothesis testing. Psychological Review, 94, 211-228.
Lehrer, R., Schauble, L.,* Carpenter, S., & Penner, D. (2000). The inter-related development of inscriptions and conceptual understanding. In P. Cobb, E. Yackel, and K. McClain (Eds.). Symbolizing and communicating in mathematics classrooms: Perspectives on discourse, tools, and instructional design. Mahwah, NJ: Erlbaum.
Mayr, E. (1982). The Growth of Biological Thought: Diversity, Evolution, and Inheritance. Cambridge, MA: Belknap Press of Harvard University Press.
Mayr, E. (1997). This is Biology: The Science of the Living World. Cambridge, MA: Belknap Press of Harvard University Press.
Mendel, G. (1959). Experiments on plant hybridization. In J. Peters (Ed.), Classic papers in genetics. New Jersey: Prentice Hall. Original publication date: 1865.
National Research Council. (1996). National science education standards. Washington DC: National Academy Press.
National Research Council. (1999). How people learn: Brain, mind, experience and school. Washington DC: National Academy Press.
O'Hara, R. J. (1988). Homage to Clio, or, toward a historical philosophy for evolutionary biology. Systematic Zoology, 37, 142-155.
Passmore, C. M., & Stewart, J. (2000). A course in evolutionary biology: Engaging students in the "practice" of evolution." Research report 00-1 for the National Center for Improving Student Learning & Achievement in Mathematics & Science. (http://ncisla.wceruw.org/publications/reports/RR00-1.PDF)
Passmore, C.M., & Stewart, J. (in press) A modeling approach to the teaching of evolution at the high school level. Journal of Research in Science Teaching.
Richards, R. J. (1992). The structure of narrative explanation in history an biology. In M. H. Nitecki & D. V Nitecki (Eds.), History and Evolution (pp. 19-53). Albany, NY: State University of New York Press.
Rudolph, J. L. & Stewart, J. H. (1998). Evolution and the nature of science: On the historical discord and its implications for education. Journal of Research in Science Teaching, 35 (10), 1069-1089.
Stewart, J., & Hafner, R. (1991). Extending the concept of "problem" in problem-solving research. Science Education, 75, 105-120.
Wynne, C., Stewart, J., & Passmore, C. (in press). High school students' use of meiosis when solving genetics problems. International Journal of Science Education.
White, B. Y., & Frederiksen, J. R. (1998). Inquiry, modeling, and metacogntion: Making science accessible to all students. Cognition and Instruction, 16, 3-118.
Wirtz, K.S. (1993). High School student conceptions of evolution: Epistemological and experimental considerations. Unpublished master's thesis, Cornell University, Ithaca, NY.
1 Discrete traits are those for which two or more distinct categories of phenotypes (or variants) are identified. For example, Mendel studied the trait of height in pea plants. He noted that the pea plants were either short (18 in.) or tall (84 in.). In contrast, height is not a discrete trait in humans: human height is best characterized as continuously variable, or nondiscrete, because humans are not simply either 18 inches or 84 inches tall. Thus, the phenotype categories for height in humans are not clear-cut.
2 Achondroplasia is inherited in a codominant fashion. Individuals with two disease alleles (2,2) are severely dwarfed and seldom survive. Individuals who are heterozygous (1,2) are achondroplastic dwarfs, having disproportionately short arm and leg bones relative to their torsos. Thus while these two phenotypes differ from normal stature, they are distinct from one another.
3 In the past, our students have developed the following explanations for protein action in traits inherited in a codominant fashion:
- One allele (designated '1') codes for an active protein. The other allele
codes for an inactive protein. Thus, individuals with genotype (1,1) have
the greatest amount (or dose) of active protein and the associated phenotype
at the organismal level. Individuals who are (2,2) have little or no measurable
protein activity and this is reflected in the phenotype. The heterozygous
individuals (1,2) have an intermediate level of protein activity and a
phenotype that is also intermediate.
For example: In the case of achondroplasia, (1,1) individuals might have two alleles for a growth receptor and a phenotype of normal stature; (2,2) individuals have little or no functional receptors and suffer from severe growth retardation; and the heterozygotes (1,2) will have half as much growth receptor activity as the (1,1) individuals and consequently are short statured achondroplastic dwarves without the additional health problems of the (2,2) individuals. This example of codominance is admittedly simplified, as students do not study the systemic effects of achondroplasia. However, this model is applied widely in genetics and sometimes referred to as the "dosage" model.
- Both alleles code for active proteins, giving rise to observable phenotypes
at the macroscopic level. Heterozygotes display the phenotypes associated
with both alleles.
For example: in human blood types, individuals carrying alleles for Protein A and Protein B have both of these proteins on their blood cells. The phenotype is not blended or dosage dependent as in the achondroplasia example above. Instead, both proteins are detected intact in the heterozygous individual.